
Useful Links
A seminal review on BNST function:
Walker et al. (2009),
A recent and comprehensive review:
Daniel & Rainnie (2016),
Our contribution for the Journal of Neuroscience Dual Perspectives series:
Gungor & Pare (2016),
And the companion paper focusing on human BNST:
Shackman & Fox (2016)
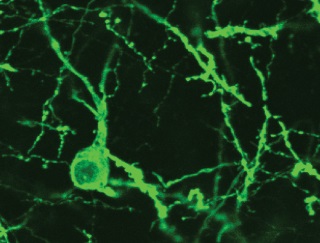
Top: A confocal image of a CeA-projecting BNST cell expresses GFP and ChR2.
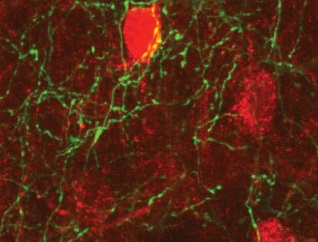
Bottom: mCherry-expressing CeA cells are contacted by GFP-expressing BNST axons.
(Modified from here)
Interactions between BNST and the central amygdala (CeA)
Early work stressed the differential involvement of the CeA and BNST in the genesis of fear versus anxiety, respectively. It was hypothesized that BNST mediates sustained anxiety-like responses to diffuse environmental threats, as opposed to the CeA, which generates defensive behaviors in response to imminent threats (Walker et al., 2009). Although this model became extremely influential, accumulating evidence suggests that BNST’s influence is not limited to the generation of anxiety-like responses but that it also shapes responses to discrete threatening stimuli. Therefore, a thorough understanding of fear/anxiety generation can only be reached by understanding BNST-CeA interactions.
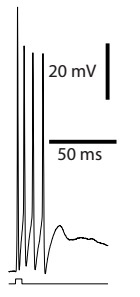
We started to study BNST-CeA interactions by characterizing the projections from BNST to CeA. It was known that most BNST cells are GABAergic except for a small population of glutamatergic cells in the ventral part of BNST (BNST-AV). We used a double viral strategy to drive expression of ChR2 in BNST cells that project to CeA. In vitro recordings showed that most of the light-evoked responses were inhibitory; revealing that the glutamatergic cell population in BNST-AV contributes little projections to CeA. This experimental paradigm also enabled us to investigate the local connectivity in BNST. Interestingly, in BNST-AV, connections from CeA-projecting to non-CeA targeting cells in BNST were distinctively low, suggesting independent processing networks may exist in BNST-AV.
A light-evoked spike burst in a ChR2-expressing BNST neuron that project to CeA.
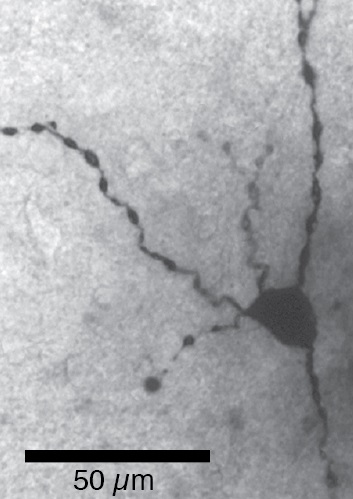
The soma and proximal dendrites of a glutamatergic cell revealed by biocytin filling.
Contrasting the properties of glutamatergic and GABAergic BNST-AV neurons
As noted above, most BNST cells are GABAergic. However, a small population of glutamatergic cells exists in BNST-AV. Importantly, there is evidence that the two cell types exert opposite influences on negative emotional states (Radley et al., 2009; Jennings et al., 2013): GABAergic cells reduce, whereas glutamatergic cells increase fear/anxiety. Intriguingly, despite being few in number, the influence of glutamatergic cells on behavior seems to dominate.
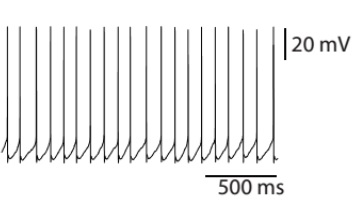
Spontaneous firing exhibited by a glutamatergic cell.
This information led us to investigate the properties of these two cell types using various techniques. Do the afferent and efferent connections of the glutamatergic and GABAergic cells differ? Are there differences in their intrinsic electrophysiological properties, morphology, or responsiveness to noradrenaline?
CGRP effects on BNST-AL neurons
BNST has direct projections to autonomic and neuroendocrine regulatory structures of the brain which are thought to evoke fear and anxiety-like behaviors. Furthermore, most BNST cells are GABAergic and are expected to inhibit their targets. In contrast with this, infusion of calcitonin gene-related peptide (CGRP) into BNST was reported to activate BNST targets and potentiate anxiety (Sink et al., 2011).
In this project, we investigated the effects of CGRP on anterolateral BNST (BNST-AL) neurons using whole-cell patch clamp recordings in vitro. We found that CGRP postsynaptically increases the inhibitory inputs evoked by electrical stimulation of the stria terminalis, one of the main input pathways to BNST. In contrast, CGRP has no effect on excitatory inputs. Thus, our findings suggest that CGRP potentiates anxiety-like behaviors and increases neural activity in BNST targets by inhibiting BNST-AL cells.
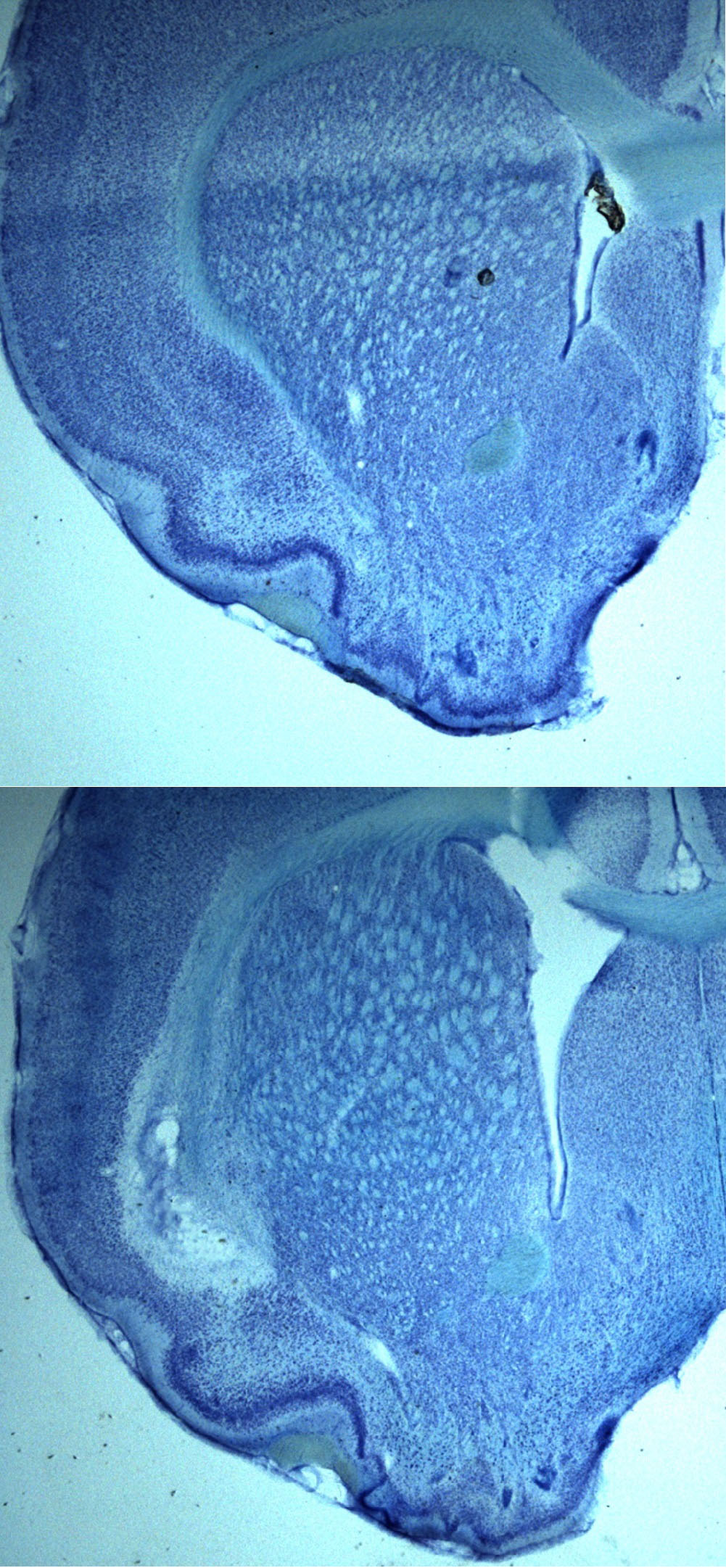
Nissl stained insular cortex sections
Top:Sham
Bottom:Ibotenic acid lesion
Interactions between the insular cortex and BNST-AL
There is ample evidence suggesting corticotrophin releasing (CRF) cells in BNST-AL are anxiogenic. However, it is unclear which structures “inform” CRF cells of threatening environmental contingencies. For instance, the oval nucleus (where CRF cells are concentrated) is devoid of inputs from basolateral amygdala, subiculum or medial amygdala, in contrast to other BNST subnuclei. However, the oval nucleus receives selective viscerosensory afferents from the anterior and middle insular cortex.
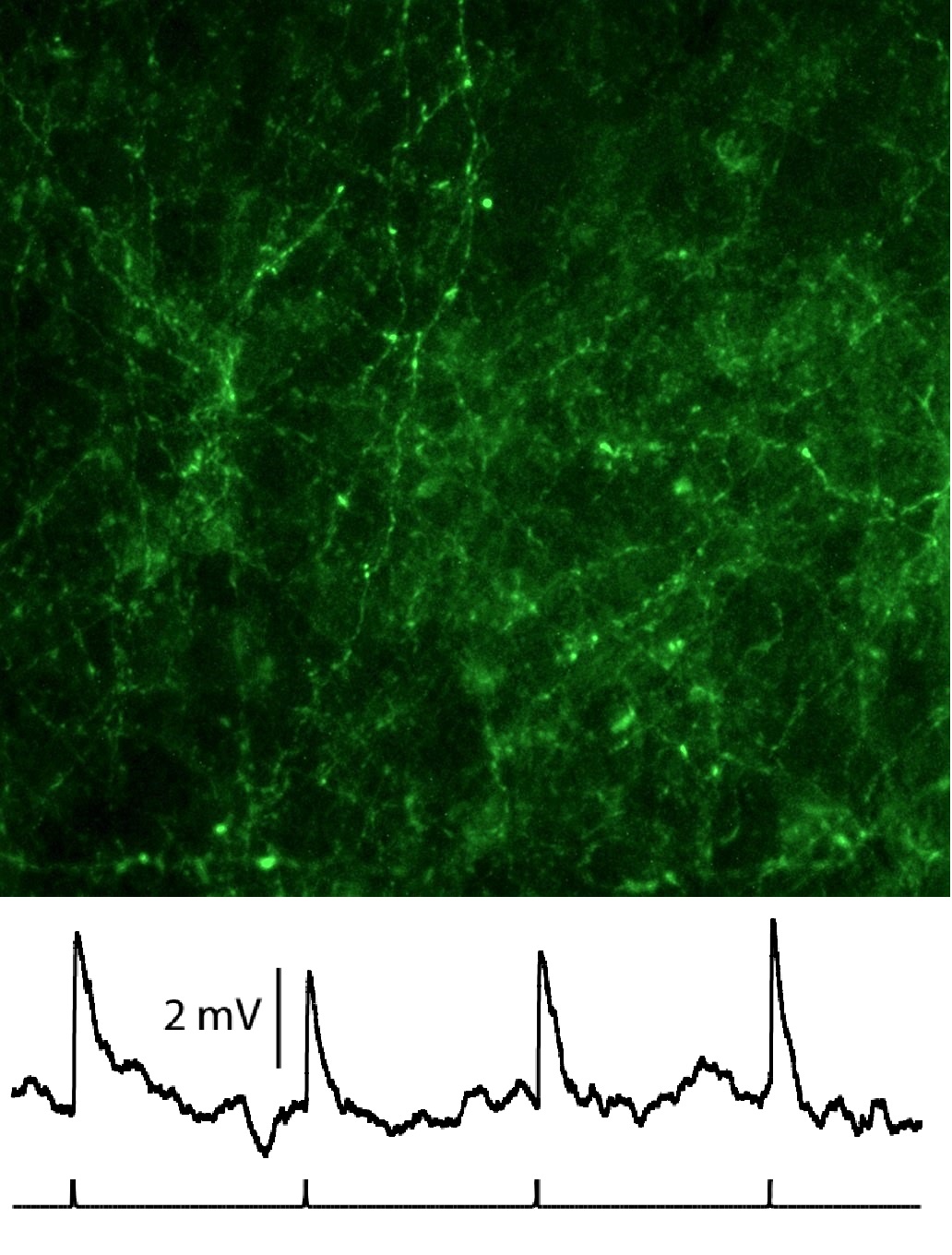
Top:ChR2 and GFP expressing insular cortex axons in BNST-AL.
Bottom: Light-evoked (1 Hz) excitatory potentials in BNST-AL.
The anterior insular cortex (in contrast to its gustatory posterior counterpart) has been implicated in a wide range of complex behaviors, as well as negative emotions (for reviews on human studies: Craig, 2009; Feinstein, 2013). Therefore, we set out to characterize the interactions between the insular cortex and BNST. First we tested the effects of anterior insular lesions in behavioral tests that are known to involve BNST activity like contextual fear conditioning and exposure to elevated plus maze. However, ibotenic lesions did not affect performance. On the other hand, there is evidence that both BNST and anterior insular cortex lesions decrease inhibitory avoidance, suggesting this behavioral paradigm is a good candidate to study insular cortex-BNST/oval nucleus interactions.